This position has been filled. See People for a list of current lab members. Feel free to get in touch for future opportunities to join our lab.
Our lab is looking for a new PhD student on a fully-funded 4-year studentship to start as early as October 2024 at Uppsala University. The PhD project will study why does buzz pollination evolve using a series of experiments in the lab and the field. If you are interested in evolutionary biology, plant-pollinator interactions, and enjoy doing experiments with live bees and plants, this project might be for you.
For more details and how to apply see below.
Deadline: 18 September 2024
The project
Buzz pollination is a type of pollination in which bees use vibrations to shake pollen out of flowers. Buzz pollination is critical for thousands of plant species as well as for crops like tomato, eggplant, and kiwifruit. Buzz pollinated flowers have evolved independently across many and different lineages , with perhaps as many as 145 independent origins across all flowering plants, making them one of the most widespread and striking examples of convergent evolution in floral form. Yet, we still do not know why plants evolve buzz pollinated flowers. The repeated evolution of buzz pollinated flowers, provides a powerful way to test hypotheses for why buzz pollination evolves, thus contributing to solve the biological puzzle of the repeated evolution of an ancient pollination system that includes thousands of bee and plant species.
The main aim of this project is to leverage the repeated evolution of buzz pollination across multiple plant lineages to test four general hypotheses of why buzz pollination evolves. The proposed project is of general significance because it addresses a fundamental phenomenon in biology: Why do similar morphological structures evolve in distantly related lineages?
We offer a fully-funded 4 year PhD studentship open to qualified applicants of all nationalities. Our lab is based at the Department of Ecology and Genetics, which is part of the Evolutionary Biology Centre (EBC) at Uppsala University. Within the Department, our lab is part of the Program in Plant Ecology and Evolution, which is home of about 8 full-time faculty, and several postdocs and PhD students. You can find more about research in our program here. Researchers and students here have access to excellent glasshouse and controlled environment facilities, as well as fully equipped molecular labs and high-performance computer clusters. The Department works closely with the national infrastructure facility SciLifeLab. In our lab, we have access to specialised equipment to study buzz pollination including a laser vibrometer, high-speed camera, particle counter, force and acceleration sensors, and other useful tools.
EBC is a dynamic and welcoming place where to conduct evolutionary studies and includes research groups working across all levels of biological organisation (from molecules to ecosystems) and study systems (plants, animals, fungi, bacteria, etc.). Our Department is highly international and we welcome researchers and students from all over the World. PhD students are involved in taking courses as well as conducting research, and there are options for gaining teaching experience as teaching assistants. At Uppsala, we also have a Biology PhD School that brings together students from multiple departments and organises symposia, courses, newsletters, and science meetings for PhD students.
Being a PhD student at Uppsala University comes with many social benefits including sickness and parental benefits, as well as Sweden’s medical care, University’s accident insurance, etc. As a PhD student here, you are considered an employee with all the benefits and responsibilities it entails. Read more here.
Informal queries can be directed to Mario Vallejo-Marin via email. Please read the ad carefully before contacting me.
Deadline: 18 September 2024.
For full application details please check the official job ad in the University’s website (link here, notice that the ad will be published on 21 August). If it is useful for preparing ahead of time, the application should include 1) a brief letter stating your research interests, and a motivation of why you want to pursue a Ph.D., your interest in this specific project, and your suitability for the position, 2) your CV including your education, research experience, and references to publications, if applicable, and 3) names and contact details of two referees (email addresses, institution, and phone numbers).
Please cite as: Vallejo-Marin M & GC Vallejo (in press). Comparison of defence buzzes in hoverflies and buzz-pollinating bees. Journal of Zoology. https://doi.org/10.1111/jzo.12857Read the paper here.
Email me if you want a PDF copy.
Hoverflies (Syrphidae) rely on pollen and nectar from flowers.
Buzz pollination is a striking form of pollination in which bees use their powerful thorax muscles—the same that power flight—to shake flowers and rapidly remove thousands of pollen grains. This summer we embarked on a quest to answer a simple question: Why flies do not buzz pollinate?[Read the full paper here].
Among the precious resources that flowers offer to their pollinators, one of the most important ones is pollen. Pollen is rich in proteins and bees and some flies rely on pollen grains to mature their sex organs, nurture their developing eggs, and provision their larvae. In the quest for pollen, bees and flies have evolved diverse morphological and behavioural adaptations to obtain and collect pollen grains, including feather-like hairs, pollen baskets and many others.
One of the behavioural adaptations evolved by pollinators is the capacity to buzz flowers to remove pollen, something that is often called sonication due to the loud sound that these floral buzzes produce. This pollination buzzes are produced by the same muscles that power flight. Typically, a buzz-pollinating bee grabs the pollen-carrying anthers with its mandibles, curls the body around the antrhers and begins rapidly contracting its thoracic muscles. The vibrations transmitted to the flower result in the ejection of thousands of pollen grains. In some specialised flowers with anthers that open only through small pores at their tip, buzzing flowers is the only way to efficiently remove pollen. Buzz pollination is relatively common in bees, having evolved in more than 11.000 species. In contrast, buzz pollination is extremely rare in flower-visiting flies, and only 1 out of 6,000 has been reported to buzz pollinate. Why is buzz pollination so rare among flies?
A hoverfly (Cheilosia illustrata) foraging in the inflorescence of an umbel plant (Apiaceae).
One of the hypotheses explaining why some insects do not buzz pollinate is that they are unable to produce vibrations of sufficient magnitude to remove pollen from flowers. Pollen release is a function of the amplitude or magnitude of the vibrations more than it is of frequecy or pitch. If flies are not capable of producing vibrations of sufficient amplitude, then there is no point in buzzing flowers. We called this the amplitude-limitation hypothesis.
A bumblebee (Bombus sp.) buzz-pollinating a flower of Solanum dulcamara (woody nightshade) in the authors’ garden.
Testing the amplitude-limitation hypothesis
To test this hypothesis we set out to compare the vibrations produced by flies and buzz-pollinating bees. We used a type of vibration that is similar to floral buzzes: Defence vibrations. Both bees and flies produce buzzes when they are threatened or alarmed. These defence buzzes are produced with the same muscles as pollination buzzes. Importantly, defence buzzes can be induced by gently squeezing the insect or simply by breathing on it (some insects respond to the CO2-rich breath of potential predators).
We measured defence vibrations using a custom-made set up that consisted of a miniature (0.2 gram) accelerometer tied to a bamboo stick. The accelerometer converts vibrations into electrical signals that can then be recorded and analysed in a computer. Our goal was to measure vibrations in as many bees and flies as possible. Limits on travel during the summer of 2020, meant that we focused our efforts in Scotland. Exploring Scotland and some of its most beautiful islands in Orkney and the Outer Hebrides was not too bad.
Looking for flies and bees in the Isle of Harris in the Outer Hebrides.
Nearly 300 insects and 4,000 buzzes later
During our experiments we measured nearly 4,000 vibrations from about 300 insects in more than 40 taxa. The hoverflies we studied included some beautiful bumblebee mimics such as the large Volucella bombylans,Merodon equestris, and Cheilosia illustrata. The exploration of the Sottish islands also allowed us to study some bee species and forms that are rare or absent from mainland Scotland such as Bombus muscorum agricolae and Bombus jonellus. We also were lucky to find a nest of the rare great yellow bumblebee (B. distinguendus) in the Isle of Harris but decided against sampling these bees as they are of conservation concern. For each defence buzz we estimated the fundamental frequency and the peak acceleration amplitude (in m/s^2). For each insect we also measured its thorax which contains the muscular apparatus powering the vibrations used by bees and flies. Our findings are simple: After accounting for insect size, we found no difference in the amplitude of defence buzzes produced by flies and buzz-pollinating bees. In other words, our results provide no support for the amplitude-limitation hypothesis.
A hoverfly, Volucella bombylans (left), and a bumblebee, Bombus jonellus (right).
Other hypotheses: Pollen needs
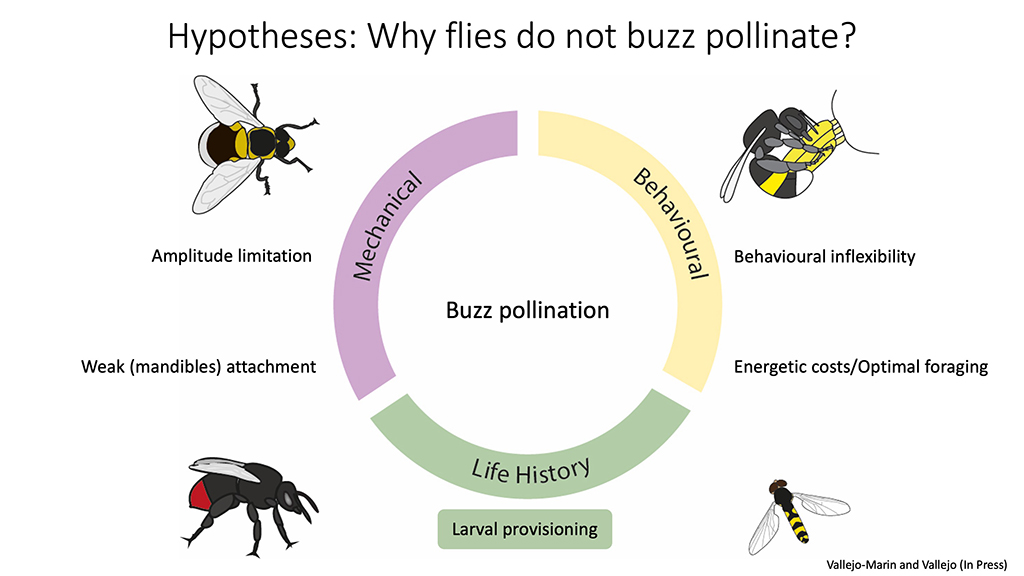
The reason for why flies do not buzz pollinate remains mysterious, but in our paper we propose some hypotheses that could explain the lack of buzz-pollinating flies. Briefly, these hypotheses can be grouped in mechanical, behavioural and life-history explanations. The amplitude-limitation hypothesis is a mechanical explanation. Another mechanical-morphological hypothesis is that the lack of mandibles in flies prevents them from holding to the flower hard enough to effectively transmit vibrations–or from becoming dislodged from the flower while buzzing! Behavioural explanations such as an inability of flies to deploy buzzes across different contexts (eg, using defence buzzes in flowers), seems unlikely given the wonderful behavioural flexibility exhibited by many flies.
Hypotheses for why flies do not buzz pollinate
Finally, life-history explanations include the possibility that flies simply do not need to collect large amounts of pollen from flowers. Unlike bees, flies consume pollen on the flower and do not need to carry it back to a nest. Bee larvae rely entirely on pollen for protein as they develop, while many overfly larvae are predatory or use other sources of organic matter. We think that this type of explanations provide the ultimate answer of why flies have so rarely evolved the capacity to buzz pollinate, but these hypothesis await testing in future experiments.
The lab has several opportunities for competitive PhD studentship applications for the upcoming academic year starting in October 2021.
This year we are offering projects on bee pollination, plant speciation, invasive species and adaptive evolution, among other themes. The projects range from the applied to the theoretical and everything in between. We are looking for excellent, motivated students that are interested in working with a diverse team of people on the broad area of plant ecology and evolution.
The application calls are for PhD studentships awarded through a competitive application funded by NERC and BBSRC. Some of these calls are open to international applicants (NERC) but please check carefully for eligibility requirements.
If you are interested in any of these projects please contact me by email or contact the lead supervisor. Informal queries are very welcome, so please don’t be shy and get in touch!
Projects arranged by lead institution.
University of Stirling
Project 1. Behavioural biology of buzz-pollinating bees and flies. DTP IAPETUS2. NERC.
Supervisors: Mario Vallejo-Marin (U Stir), Susan Healy (U. St Andrews), Gema Martin-Ordas (U Stir).
Brief overview: More than 15,000 species of bees are capable of deploying powerful vibrations of their thorax to remove pollen from flowers with specialised morphologies. Pollination by these vibration-producing bees has given rise to the phenomenon of buzz pollination, which occurs in more than 20,000 species of flowering plants and includes important crops such as tomato, blueberries and kiwifruit. Previous work has shown that the use of vibrations to remove pollen from flowers is part of the behavioural repertoire available to some species of pollen-collecting insects, particularly bees. This project addresses two main questions (1) Why is buzz pollination is mostly restricted to a subset of bee species? (2) What determines when a bee capable of buzz pollination deploys vibrations to extract pollen from flowers?
Project 2. Bees and buzz pollination in crops: Evaluating the potential for matching bee vibrations and buzz-pollinated crops to improve fruit yield. EastBio. BBSRC.
Brief overview: Buzz-pollinated plants include important food crops such as tomato, blueberries, and kiwi. Buzz-pollinated crops grown in greenhouses or outdoor settings often require supplemental pollination to achieve full fruit set and to improve fruit quality. This supplemental pollination is often provided using mechanical shakers, or additional pollinators such as honeybees or bumblebees. However, the effectiveness of these treatments is enormously varied even within the same crop species. This diversity of outcomes could be explained by variation at the plant, bee or environmental levels. Previous work has shown that buzz-pollinating bees differ widely in the type of vibrations they can produce, and that the type of vibration applied to a flower determines pollen release, and potentially fruit quality. Moreover, environmental variables such as temperature and humidity may influence both the likelihood of bees producing vibrations or the effectiveness of these vibrations to release pollen from different types of flowers. The main goal of this project is to investigate what type of supplemental pollination is more effective at maximising fruit yield in buzz-pollinated soft fruits including blueberries. In this project, we will use laboratory and field experiments to address three questions: (1) What is the relationship between vibration properties (amplitude, frequency and duration) and pollen release and fruit quality across different varieties of blueberries? (2) What are the properties of the vibrations used by buzz-pollinating bees while visiting experimental plots of different varieties of blueberries? (3) What is the effect of increased temperature and humidity on the capacity of buzz-pollinating bees to release pollen from flowers?
On the road: Range expansion of coastal plants under global change
Supervisors: Dan Chapman (UStir), James Bullock (CEH), Kevin Walker (CEH) and Mario Vallejo-Marin.
To cope with human-driven environmental change species must either move or adapt. Indeed, shifts in the geographical distributions of species are a clear ecological response to environmental change. Global climate change is driving poleward and uphill range expansions in many species, while other species exploit anthropogenic habitat modification and human-mediated dispersal to colonise new habitats. Better understanding of this range-shifting ability is needed to predict ecological responses to environmental change. This project will achieve that by identifying the species’ characteristics that are associated with successful range expansion.
The studentship will focus on range expansions by coastal specialist plants, which have been understudied compared to fully terrestrial species. Coastal salt tolerant plants (termed halophytes) are a tractable group to investigate multiple dimensions of range expansion in a way not possible for other species groups. This is because coastal halophytes have two potential routes for range shifting. First, climate change could allow them to colonise further north along the coastline. Second, winter de-icing of roads using salt has created new halophytic habitats, allowing some species to spread inland onto road networks, often using vehicles for rapid dispersal. This study will be the first to systematically investigate and compare range expansions into natural vs anthropogenic territory.
Brief overview: This project will analyse genome data from UK populations of Mimulus guttatus (yellow monkeyflower) to determine how the local ecology and geography affects its reproductive mode.
The project will involve the analysis of new genomic data from previously–collected samples to quantify the extent of self-fertilisation and clonal reproduction in natural populations in the UK. This data can then be compared with geographical, ecological, and climatic data to determine if these factors correlate with the local reproductive mode. Genetic data from UK samples will then be compared with those from the United States, to determine their population history and investigate if different degrees of asexual and sexual reproduction arose at specific times in the past. These data will allow us to investigate how the evolutionary history differs between populations exhibiting different reproductive modes. There will also be scope to develop theoretical models to further investigate the consequences of reproductive mode on genetic variation, or to collect more samples for analysis. The project will provide the student with cutting-edge genomics and bioinformatics skills, which are essential for modern biological research, and put them at the forefront of developing an exciting new model for evolutionary genetics study.
Brief overview: Rhododendron ponticum is invasively naturalised over much of the British Isles, which is much colder than its native range (Spain and Turkey). Naturalised material contains introgressed (transferred by hybridisation) DNA from the highly cold-tolerant American R. catawbiense, and such DNA is more frequent in colder regions of the UK, suggesting but not proving that introgression from R. catawbiense confers increased cold tolerance.
This project would make use of the full genome sequence of R. ponticum, recently generated by co-supervisor Dr Jianquan Liu’s group in China. This will be used as a basis for designing a set of RNA baits to capture sequence data from hybrid populations and herbarium samples. The bait set will comprise candidate cold tolerant genes, plus single copy genes well dispersed over the genome, to allow for FST scanning to detect frequency of introgressed regions from R.catawbiense, and to determine if cold-tolerance associated genes are over-represented in the introgressed set.
Brief overview: Buzz pollinated plants require buzzing bees for their effective pollination. This project explores whether different floral morphologies have evolved in response to different buzz pollinating bees.
Buzz pollination has evolved among thousands of flowering plants, including species of agricultural importance such as the tomato. Buzz pollination is a specialised form of pollination in which bees extract pollen from flowers that have specialised poricidal anthers that unlock only under vibrations produced by bees.
Buzz pollinated plants rely on buzzing bees for their pollination. Yet we know relatively little about the interaction of buzz pollinating bees and different floral morphologies. In the face of worldwide declines in bee diversity, this project aims to explore how different floral morphologies evolve in response to different types of bees.
The project will explore the diversity of floral traits across buzz pollinated plants in the context of what we know about the biomechanics of buzz pollination, buzzing behaviour in bees, and other aspects of bee behaviour and diversity. For example, bees are known to prefer to visit darker coloured flowers in colder temperatures, which may in turn affect floral morphology and colouration in a given climate. Such hypothesis can be tested using georeferenced occurrence data of buzz pollinated plants, their corolla colouration, and the bioclimate of each occurrence point. There is also space within the project for the student to develop experiments using plants and bees.
The work will be undertaken in a cross-disciplinary research team between the Royal Botanic Garden Edinburgh (plant evolution & taxonomy), University of Edinburgh (ecology), and University of Stirling (plant reproduction, plant & bee interactions, buzz pollination).
Brief overview: Invasive populations are ideal model systems to understand the early stages of allopatric speciation, a keystone concept of evolutionary biology (1). Divergent selection to different environments can drive to reproductive isolation and, eventually, speciation (2). The annual, Centaurea solstitialis, (yellow starthistle) is an ideal system to study selection and speciation during a biological invasion. It is native from Eurasia, and invasive in Australia and the Americas (3). It developed different sets of local adaptations to different regions, including some degree of reproductive isolation (1,3-6). The fundamental research undertaken in this PhD project, will address the emergence and maintenance of reproductive isolation in yellow starthistle, and investigate how the balance of selective or neutral processes can lead to incipient speciation in a new allopatric range. This research will contribute substantially to our understanding of how biodiversity is generated and maintained , as well as establishing the likely impacts of invasive populations on biodiversity.